MeRIP-seqを用いて魚類のメチル化環境RNAにおける温度関連の変動を検出する
https://www.biorxiv.org/content/10.1101/2025.11.03.686443v1.full
はじめに
環境DNAは生物の存在確認や分布把握に広く使われていますが、近年は環境RNA(eRNA)も注目されています。
eRNAは生きた細胞に由来し、環境中での分解も比較的早いとされており、環境DNAよりもその時点に近い生物の状態を反映しやすいと考えられています(Cristescu, 2019; Yates et al., 2021; Veilleux et al., 2021)。特に魚類では、水中から回収したeRNAによって、生物の存在だけでなく生理状態やストレス応答まで捉えられる可能性が示されて来ました(Giroux et al., 2022; Littlefair et al., 2022; Miyata et al., 2021, Miyata et al., 2025)。
ただし、これまでのeRNA研究の多くは、主にメッセンジャーRNA (mRNA) 量の違いをみる発現解析にとどまっていました。RNAの機能は発現量だけで決まるわけではなく、安定性や翻訳効率などを左右する転写後制御も重要です。
その代表例がN6-メチルアデノシン(m6A)で、m6Aは真核生物のRNAに広く見られる修飾であり、温度変化のような環境ストレスに応じて変動することが知られています(Zaccara et al., 2019; Ahi & Singh, 2024; Wang et al., 2025)。
著者らは、eRNA研究を発現量比較からさらに進め、環境中に残るRNA修飾まえ視野に入れる必要があると位置づけています(Ahi & Schenekar, 2025)。
そこで本研究では、アフリカンシクリッド Labeotropheus trewavasae を用い、26℃ の対照区と 29℃ の高温区の水槽水から回収した eRNAにMeRIP-seqを適用し、m6A修飾情報が温度応答の把握に使えるかを検証しました。
環境中に放出されたextra-organismal RNAから、m6Aというエピトランスクリプトーム情報を直接読み取ろうとした点がこの論文の新しさであり、水試料だけで魚類の温度ストレスを評価する新しい手がかりになる可能性があります。実際に著者らは、この研究を「eRNA を mRNA Abundanceの先へ進める試み」と位置づけています。
メソッド
アフリカンシクリッド L.trewavasae を用い、高温条件が水中eRNAに与える影響を調べています。実験はと解析は、飼育条件の設定、水試料からのeRNA回収、m6Aを標的としたMeRIP-マッピングと統計解析の順で進められています。
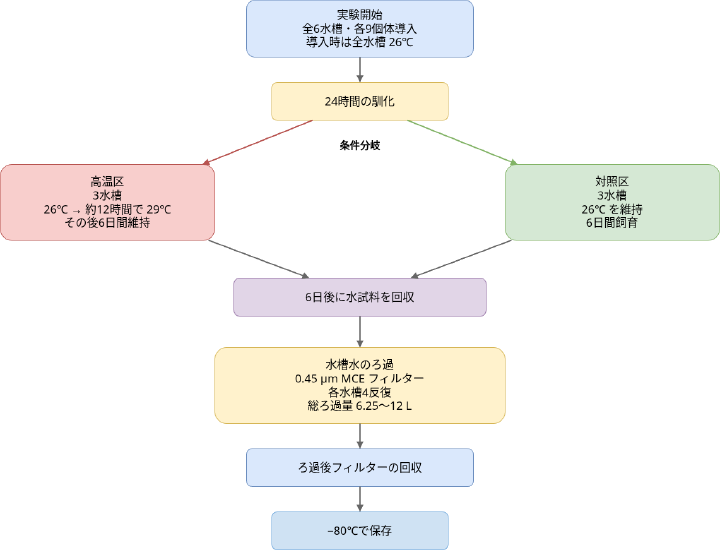
実験条件と eRNA 回収
まず、60L 水槽を6台を用意し、各水槽に L. trewavasaeを 9個体ずつ飼育した。飼育水槽の条件とその後のろ過・抽出の操作は下記の通り。
RNA 抽出と MeRIP-seq
回収したフィルターからはTrizolを用いて全RNAを抽出した。RNA収量を確保するため、各水槽の4反復のうち2反復ずつを最終段階でプールし、1水槽あたり2試料、合計12試料を作成している。抽出RNAにはDNase I処理とrRNA除去を施し、その後にRNAを断片化した。続いて、抗m6A抗体を用いてm6Aを含むRNA断片を濃縮し、MeRIP-seqライブラリを調製してIllumina PE150でシーケンスしています。
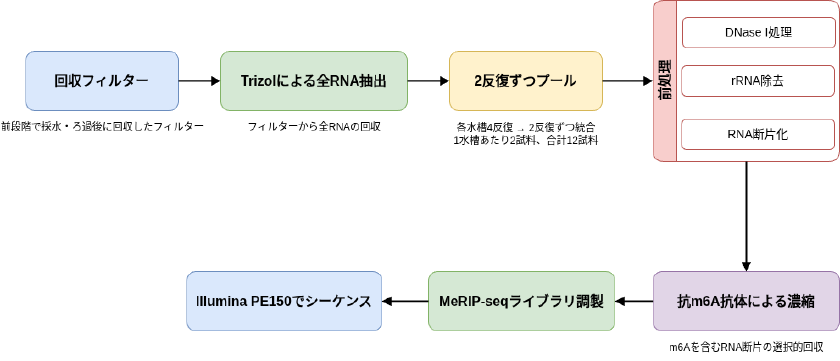
上記の処理よりこの研究では、通常のRNA-seqではなく、m6A修飾RNAを選択的に拾う手法を環境RNAに適用しています。
マッピングと統計解析
得られたリードは、まず FastQC で品質確認を行い、Trimmomatic v0.39 でアダプター除去と品質トリミングを行った。その後、FastQ Screen と Bowtie2 を用いて、対象魚 L. trewavasae 参照ゲノムに対するマッピング率を評価しています。
さらに下流の機能解析では、抽出したリードを近縁で注釈の充実した Oreochromis niloticus のゲノムへ再マッピングし、properly paired かつ uniquely mapped なリードのみを採用しています。
統計解析では、高温区と対照区の差を Wilcoxon rank-sum test で評価し、マッピング率とろ過水量・魚バイオマスとの関連を Spearman の順位相関で調べた。差次的 abundance 解析には metaseqR2 を用い、padj < 0.05 を有意基準とした。あわせて PCA、ヒートマップ、ボルケーノプロットも作成し、試料間の全体傾向を確認しています。

リザルト
まず著者らは、MeRIP-seqを適用した水由来eRNAから、対象魚L.trewavasaeに由来するリードを十分に回収できるかを確認しています。そのうえで、温度条件間で差のあるeRNAを抽出し、それらがどのような機能群に偏っているかを評価しています。
結果として、マッピング率は比較的高く安定しており、温度条件によって増減する m6A濃縮 eRNAが多数検出されたことが、本研究の中心的な結果になっています。
L. trewavasaeへの MeRIP-eRNAリードのマッピング
MeRIP-eRNAライブラリから得られたリードのうち、どの程度が対象魚の参照ゲノムにマップしたかという点について下記にまとめました。
全12ライブラリにおいて、クオリティフィルタリング後のリードのうち平均 22.6%が L.trewavasaeの参照配列にマップし、サンプルごとに見ると13.6〜29.4%の範囲でした。高温区の平均は 24.2%、対照区の平均は 21.0%と、高温区の方がやや高いものの、統計的には有意差は認められていません。
したがって、少なくとも本実験条件では、温度処理そのものが宿主由来リードの回収率を大きく変えたわけではなかったと言えます。
- 1 サンプルあたり平均 5.82億リードの取得
- L. trewavasae 参照ゲノムへの平均 22.6%のマッピング率
- 平均値だと高温区 24.2%、対照区 21.0%のマッピング率
- 温度条件間での有意差なし(Wilcoxon rank-sum test, p = 0.31)
この結果は、環境由来試料でありながら、MeRIP-seq後の eRNAから相当量の宿主シグナルを確保できたことを示しています。
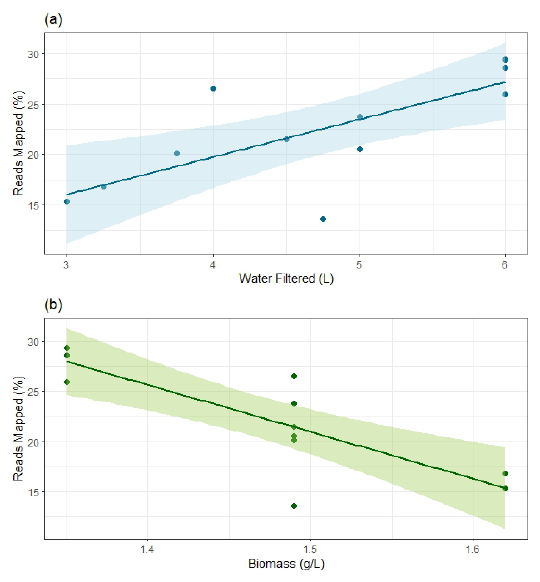
マッピング率に影響した要因
次に著者らは、宿主由来リードの割合に影響する要因を調べていまする。Fig.1によると、マッピング率はろ過水量が多いほど高く、逆に魚のバイオマスが大きいほど低い傾向を示しています。Spearmanの順位相関では、ろ過水量との間に正の相関、バイオマスとの間に負の相関が確認されています。
つまり、より多くの水をろ過した方が対象魚由来 eRNAを回収しやすく、一方で高密度飼育に近い条件では、非標的RNAの割合が相対的に増えて宿主マッピング率が下がった可能性が示唆されます。
高温区で増加したeRNA
温度条件間の差次的Abundance解析では、検出された 755の eRNAのうち、106が高温区で有意に増加していました。これらの log2FCは 0.467〜4.213で、中央値は 1.085でした。すなわちおおよそ 2倍前後の増加に相当します。
その中で、klf4, abca12, capn9, lmo7a, cdc42bpb などが注目遺伝子として挙げられていました。
GO解析では、増加した遺伝子群がintermediate filament-based process, antigen processing and presentation, cell-cell adhesion に有意に偏っていた。著者らはこれを、上皮バリア、細胞接着、細胞骨格再編成、免疫応答といった、温度ストレスに関わる生理機能と整合的なパターンとして解釈している。
- 高温区で増加した 106のeRNA
- log2 fold change 0.467〜4.213
- 主な候補としての klf4, abca12, capn9, lmo7a
- GO 解析
ブログ本文では、この結果を「高温条件では、上皮の構造維持や細胞間結合、免疫関連に関わる eRNA が増えていた」とまとめると伝わりやすい。RNA 修飾そのもののピーク差を直接論じる前段階として、m6A 濃縮分画の中で温度応答らしい機能群が立ち上がっていることが読み取れる。
高温区で減少した eRNA
一方で、306 本の転写産物は高温区で有意に減少していた。こちらは増加群よりも数が多く、log2 fold change は −0.689〜−6.700、中央値は −2.069 で、4 倍前後の低下に相当する。主な候補として mmp9, cebpb, antxr2a, socs3a, ldha, eno1a などが挙げられている。
GO解析では、pyridine-containing compound metabolic process, small GTPase-mediated signal transduction, purine-containing compound metabolic processが有意に濃縮していた。著者らはこれを、代謝系、とくに解糖系やペントースリン酸経路(PPP)やヌクレオチド代謝、さらに GTPase依存的なシグナル伝達や細胞骨格制御の抑制と関係づけています。
この減少群は、増加群よりも規模が大きく、高温条件下で代謝やシグナル伝達の一部が抑えられている可能性を示している。結果としては、高温で上がる群と下がる群の両方があり、その機能的な偏りが温度ストレス生理と対応している可能性がある。
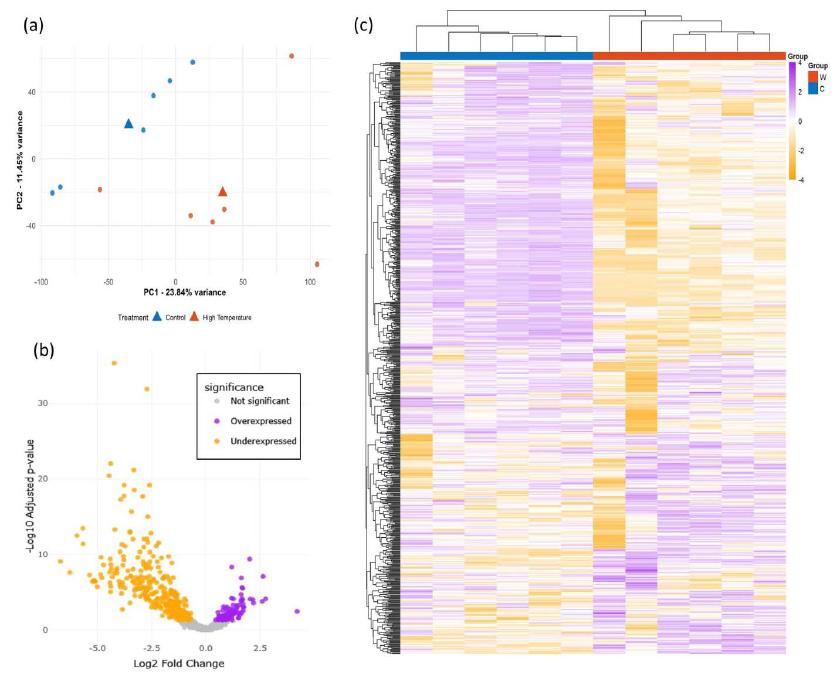
試料全体の分離パターン
試料全体の傾向は Fig.2にまとめられています。PCAでは温度条件ごとのまとまりが可視化され、ボルケーノプロットでは有意差のある転写産物が両方向に分布していることが示されている。
さらにヒートマップでは、755の転写産物について、対照区と高温区で発現パターンに違いがあることが確認できます。これらの図は、個々の遺伝子変化だけでなく、試料全体として温度条件に応じたプロファイルの差があることを示唆しています。
ここまでをまとめると、この論文では、以下が着目すべき結果になります。
- 水中 eRNAを使って宿主由来の m6A濃縮 RNAを安定して取得できたこと
- そして温度上昇に応じて増減する eRNA群が検出され、その機能が上皮・免疫・代謝応答と関係していたこと
考察
著者らは、今回の結果を単なる実験成功例としてではなく、環境RNA研究を「転写産物量の比較」から「エピトランスクリプトーム情報の取得」へ広げる最初の一歩として位置づけています。
結果から言えること
著者らは、水槽水から回収したextra-organismal eRNAに対して、通常の抗体ベースMeRIP-seqを適用し、宿主由来のシグナルを十分に得られた点が着目すべき点であるとしています。
平均で約23%というマッピング率は、従来のマクロ生物を対象としたeRNAのmRNA-seq研究よりかなり高く、m6Aを標的にしたことで真核生物由来RNAを相対的に拾いやすくなったのではと考察しており、著者らはこの点を、環境試料で問題になりやすい微生物RNAの過剰混入に対する、ひとつの実用的な打開策として見ています。
さらに重要なのは、温度上昇に応じて増減したeRNA群が、単なる無作為な変化ではなく、熱ストレス生理と関係づけやすい機能群に偏っていた点です。
高温区で増加した転写産物群は、上皮バリア、細胞接着、中間径フィラメント、抗原提示といった機能に結びつき、高温区で減少した群は、解糖系やペントースリン酸経路、ヌクレオチド代謝、小型GTPaseシグナル伝達などに関わっていました。
著者らはこれを、高温下で上皮組織の保全や再編成が進む一方、代謝やシグナル伝達の一部が抑制されるという、魚類の温度応答として解釈しています。
加えて、増減した、klf4,foxo3,slc1a5,ldha,mmp9,socs3などは、脊椎動物でm6A制御との関連がすでに報告されており、今回の研究ではm6Aピーク差そのものを頑健に議論するには限界があるものの、少なくとも温度応答に関わるeRNA群がm6A濃縮分画の中で観察されたことは、環境中に残るRNA修飾情報が生理状態の指標になりうることを示しています。
方法上の限界
一方で、この手法における限界も比較的明確となっており、最大の制約は、リプリケート数と入力 RNA量の少なさです。
今回の実験では、水槽ごとに得られた環境RNAをプールしながら12試料を解析しているが、それでもピークレベルでの差次的m6A解析を行うには十分な検出力が得られていません。そのため、今回しっかり示せたのは 「m6A濃縮 RNA群に温度関連の差が見られたこと」 であって、 「個々の m6A修飾部位が温度でどう変わったか」 を決定的に示せたわけではありません。
また、MeRIP-seq自体が持つ一般的な注意点もあります。抗体ベースの m6A解析では、抗体特異性やピークコールの再現性が問題になりうることが以前から指摘されており、環境試料のように RNA量が少なく断片化も進みやすい試料では、その影響がさらに大きくなえます。著者らも、今回の結果を方法論的なproof-of-conceptと位置づけつつ、厳密なベンチマークの必要性を認めています。
さらに、実験条件に関する限界も示されており、今回の結果では、マッピング率がろ過水量に正に相関し、魚のバイオマスに負の相関を示しました。
これは、同じ処理条件でも採水量や飼育密度の違いによって宿主由来 RNAの割合が変わりうることを意味しており、今後この手法を比較実験や野外応用に広げるには、ろ過量やバイオマスのような共変量を厳密に管理する必要がります。
今後の展望
それでも本研究の意義は大きく、著者らは、今回の結果を環境 RNA研究を「mRNA abundance の先」へ進める最初の実証例として捉えています。
技術的には、DART-seq, MAZTER-seq, m6A-SAC-seq, miCLIP2, NanoporeのDirect RNA-seqなど、より低inputまたは抗体非依存の方法も言及しており、これらは、input RNA が少ない環境試料に対しても、より高い感度や塩基レベルの分解能を与える可能性があります。
したがって次の段階としては、rRNA除去の最適化, 低input試料向け m6A手法の導入, ピークレベルでの再現性評価, 野外試料での検証が重要になるといえます。
まとめ
この研究は、魚が水中へ放出した環境RNAから、m6Aを含むRNA分画を回収し、温度条件に応じた変化を検出できる可能性を示しました。実際に、高温区と対照区の間で差次的abundanceを示すeRNAが412種検出され、その機能は上皮バリア、細胞接着、免疫応答、代謝調節など、温度ストレスと関わりの深い領域に偏っていました。
一方で、本研究はproof-of-conceptの段階であり、m6A修飾部位そのものの変化を強く結論づけるには限界もあります。とはいえ、環境中に放出されたRNAがエピトランスクリプトーム情報を保持しうることを示した点は新しく、環境RNA研究を発現量比較からさらに一歩進める結果といえます。